Chapter 13. THE REAL UNICORN ELASMOTHERIUM
It has been established that the myth of the unicorn of justice originated in China under the influence of the ancient Eurasian legal paradigm of animal judgements. But this still does not explain why people believed so fervently in the strange idea of an animal with a single horn in the middle of its forehead. For, however remote in space and time, even the most ancient of myths is usually based on some kind of reality. Therefore, is it possible that a real unicorn once roamed the earth? And could a lingering memory of such an extraordinary beast have survived, and perhaps provided the basis for the Chinese belief in the mythic unicorn zhi? Incredible as it may seem, the answer to each of these questions is a resounding “YES,” for between 100,000 and 10,000 years ago, a true unicorn called the Elasmotherium flourished on the Eurasian continent.
It is largely due to the brilliant work of Björn Kurtén that it is now possible to arrive at some kind of understanding of the great beast Elasmotherium. Most essential for my research on the Elasmotherium have been his three books, Pleistocene Mammals of Europe (1968), The Age of Mammals (1971) and The Ice Age (1972).5 However, the earliest Western scholar to identify the giant unicorn Elasmotherium was Johannes Fridericus Brandt (1802–1879) who published his classic unicorn treatise in St. Petersburg, in 1878. Unfortunately it is written in a highly technical style of Latin and deals almost exclusively with the enormous teeth of the beast (fig. 25).

Figure 25. Drawing of the molar of an Elasmotherium
In a sense, this narrow focus is understandable, because although some giant mammals carried defensive horns, the real secret of their success was the development of highly efficient teeth for grazing. Due to the enormous size of its molars, Chinese paleontologists call the Elasmotherium the ban chi xi 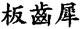 or “Plank-teethed Rhinoceros.” Apparently its huge grinding teeth are as big as planks of wood! Truly a gigantic animal, it was even larger than a modern male elephant. Its head alone measured 95 centimetres from the tip of its nose to the back of its skull (fig. 26).
or “Plank-teethed Rhinoceros.” Apparently its huge grinding teeth are as big as planks of wood! Truly a gigantic animal, it was even larger than a modern male elephant. Its head alone measured 95 centimetres from the tip of its nose to the back of its skull (fig. 26).
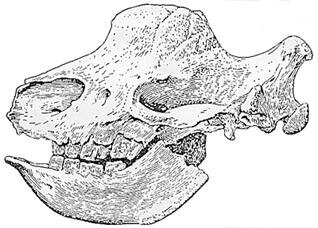
Figure 26. Drawing of the skull and mandible of the giant unicorn Elasmotherium
Most intriguing for this study is the fact that it carried a single horn which grew from the centre of its forehead and measured up to two metres in length!
But what kind of animal was this giant unicorn? Where did it come from, and what did it look like? In order to solve these mysteries we must journey back in time to the end of the Cretaceous period when large herds of great horned dinosaurs, such as Triceratops, roamed the earth (fig. 27).
Period Age in millions Dominant forms of life of years ago
Jurassic 140-180 Dinosaurs, Brontosaurus 
Cretaceous 63-140 Dinosaurs, Triceratops 
Tertiary 3-63 Mammals, birds, Indricotherium 
Quaternary Today – 3 Humans, ape-men 
Figure 27. Chart of the History of the Earth
Unfortunately, our abiding fascination with these ancient reptiles has obscured the fact that during the following Tertiary era, gigantic mammals evolved which were to surpass all but the largest of the dinosaurs in size (Kurtén, 1971, 28).
Sixty-five million years ago the extinction of the dinosaurs marked the dawn of the Age of the Great Mammals, including the Elasmotherium. The term therium (plural theria) which forms part of the Latin or taxonomical designation for this huge archaic mammal comes from the Greek therion meaning ‘a wild animal’ or ‘a beast.’ Thus the name of the great unicorn is composed of the Greek word elasma (genitive elasmos) meaning ‘a thin plate’ and therium meaning ‘mammal’ or ‘beast.’ All mammals, including human beings, belong to the zoological class Mammalia. Mammals are then grouped into orders, families, and genera in an effort to clarify the evolutionary relationships of the various species (Kurtén, 1968, 31).
As a result of the process of adaptive radiation, which is “the evolution of diversified descendants out of a single ancestral stock” (Kurtén, 1971, 30), huge mammals such as the Elasmotherium were to flourish on earth for a span of 60 million years. The majority of these great beasts would finally succumb to the increasing glaciation of the earth’s surface during the Pleistocene Ice Age (map 3).

Map 3. Greatest Extension of Continental Ice Sheets in the Pleistocene Period
However, there are certain areas of the world, known as “refugia” or “places of refuge” which were never covered by the vast continental ice sheets. There, a few huge archaic species survived down into late Pleistocene times before they were probably brought to extinction by human hunters. One of the largest of these refugia, the East Asian Refugia (map 4),

Map 4. Map of the East Asian Refugia
encompassed the steppes and forests of southeastern Russia, Mongolia and northern China, and it was there that the giant unicorn Elasmotherium lived and died, leaving fossil traces of its passing.
Recent research by B. Su and L. Jin indicates that a hominid fossil gap still exists in China between 100,000 and 40,000 years ago with no specimens found to date. However, there is fossil evidence to indicate that there were archaic humans in the area of Gansu province between 38 and 39 thousand years ago. Later, a group of people migrating up from the south, “reached the Upper Yellow River basin about 30,000 years ago, where they created one of the earliest Chinese cultures and eventually migrated to the Himalayan plateau about 6,000 years ago” (Su and Jin, 2001, 120-121). Thus early human beings living in the area of what is now north China and Mongolia could have had contact with the Elasmotherium for more than 25,000 years, an estimate which may eventually be clarified by modern genetic and archaeological research. A personal encounter with such an awesome beast would certainly have left a lasting impression, and given rise to stories about the great unicorn told around evening campfires for thousands of years to come. Gradually memories became legend, and legend faded into myth.
The oracle bone character gu (fig.28) meaning ‘ancient’ clearly reflects this long oral tradition.

Figure 28. Oracle bone and modern characters gu (ancient)
It is composed of the character for ‘shield’ (indicating war) above the character for ‘mouth’ (indicating spoken stories) and expresses the idea of war stories (of the past) or simply ‘the past’ or ‘ancient.’ The continuing vitality of the oral tradition in China is truly astounding, for even to this day gifted storytellers are held in the highest esteem.
The full scientific designation in Latin for the real unicorn is Elasmotherium sibiricum Fischer. Yet in spite of the use of the term sibericum, the homeland of the Elasmotherium was not Siberia but southern Russia (Kurtén 1968, 41), and its range was the steppes and forests of northern Eurasia. The Elasmotherium was a species of the rhinoceros family of the mammalian order Perissodactyla, which includes all three- or odd-toed plant-eating mammals. “The name odd-toed ungulates does not necessarily mean that the number of toes is odd, although this is the rule in most groups. It means that the axis of the foot always coincides with the third or central toe, which is the strongest” (Kurtén 1968, 139).
“During the early Quaternary, there were four families [of Perissodactyla]: horses, rhinos, tapirs and chalicotheres living in China. Only the horses maintain their existence; chalicotheres became extinct in the Early Pleistocene, and the other two kinds, rhinos and tapirs, retreated gradually southward beyond China” (Xue and Zhang, 1991, 316). Today we are still familiar with the first three of these families: Equidae (horses), Rhinocerotidae (rhinos) and Tapiridae (tapirs), but the fascinating Chalicotheriidae (chalicotheres), which were giant horse-like animals with claws instead of hooves, can only be imagined (fig. 29).

Figure 29. Drawing of a Chalicothere Moropus from the Miocene of Asia and North America
“They flourished in the Tertiary but became extinct in Europe and North America before the end of that period. A few specimens of Nestoritherium show that chalicotheres survived in China until the Middle Pleistocene” (ibid., 318). “The last kind of rhino in China to be mentioned is the ‘Giant unicorn’ Elasmotherium. It is very big, much larger than any living rhino. Its single immense horn is on the forehead rather than the nose bone as in other rhinos. Its cheeks [sic ‘teeth’] are high crowned and prismatic in shape with wrinkled enamel patterns on the crown surface.” (ibid., 317)
The question is, where on earth did such an amazing animal come from, and how did it evolve? In order to discover its origins, we must turn our attention away from Asia for the moment, and look eastward across the Bering Strait to North America (map 5).

Map 5. The Bering land bridge
For it was there, during the Eocene epoch, 50 million years ago, that the family of Perissodactyla first appeared (fig. 30).
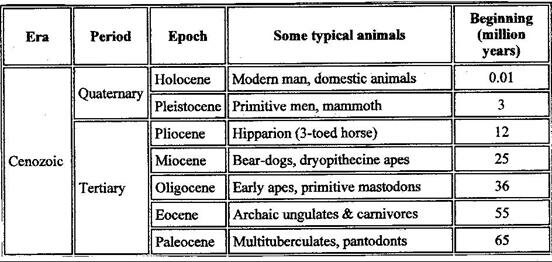
Figure 30. Chart of Geological Time and the Fossil Record
Although there is as yet no definitive proof, evidence suggests that sometime between 50 and 45 million years ago these ancient precursors of the modern rhino, horse and tapir spread westwards across Beringia, the ancient land bridge that linked North America to Asia during the Paleocene and early Eocene periods. Judging from the numbers of faunal remains of browsing animals, it is clear that the Perissodactyla flourished in the Gobi Desert area of Mongolia, which was covered by luxuriant forests at that time (Kurtén, 1971, 76).
The dispersal (westward) across Beringia did not necessarily take the form of a massive advance…. Animals living in adjoining parts of Alaska and Siberia would gradually spread into those parts of Beringia offering them suitable living conditions; if such conditions prevailed throughout Beringia, the spread might continue until the species had established a bridgehead on the new continent. This is what is actually meant when we speak of Beringia as an intercontinental road. (Kurtén 1972, 138)
It is equally possible, however, that the Perissodactyla could also have dispersed eastwards from North America to Europe across the North Atlantic land bridge. In 1972, on the basis of geophysical reconstructions and faunal similarities, the DeGeer dispersal route was proposed. This was a region of faunal interchange across the North Atlantic linking Canada with Greenland and Scandinavia until the end of the early Eocene (MacFadden, 1992, 154). This ancient land connection is now no longer a theory but a geological fact.
Approximately 45 million years ago, during the middle Eocene period, the Perissodactyla moved again, this time over the Tura Land Bridge across the Turgai Strait which linked the ancient Tethys Sea with the Arctic Ocean (map 6).
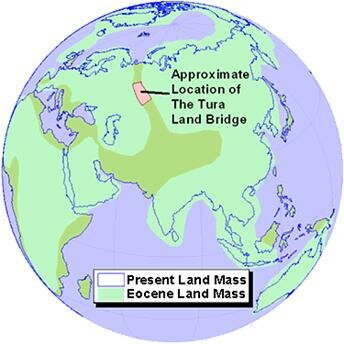
Map 6. The Tura Land Bridge
This land bridge appeared just west of the Ural Mountains, briefly joining the continents of Asia and Europe (Kurtén, 1971, 76). In Europe the rhinoceros family flourished, reaching its zenith in the late Oligocene period. The splendid prehistoric cave paintings recently discovered at Chauvet in France, which include 53 images of rhinos, provide indisputable evidence for the prevalence of the rhinoceros in ancient Europe (Chauvet, 1996). It is interesting to note that the colour scheme used in these ancient cave paintings is black, red, and white.
Evidence also suggests that during this same period of time, about 35 to 15 million years ago, a hornless giant rhino called the Indricotherium (fig. 31), which was a treetop browser much like the giraffe, roamed the area of northeastern Asia (Kurtén, 1971, 92).
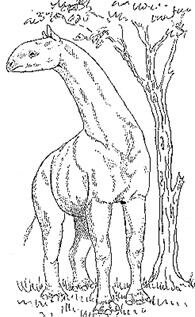
Figure 31. Drawing of an Indricotherium
For a long time the Indricotherium was considered to be the largest mammal ever to roam the earth. However, recent scholarship by Fortelius and Kappelman has revealed that this erroneous conception was based on inaccurate and inflated data.
The Oligocene giant rhinocerotoid Indricotherium transsouralicum (= Baluchitherium grangeri), ‘the largest mammal that ever lived,’ was smaller than generally believed. Over 90 estimates based on head-and-body length, skull size, molar length and proximal limb bone diameters agree well on a mean mass estimate of about 11 tonnes (t) rather than the 20-30 t given by most texts. A maximum mass estimate between 15 and 20 t seems probable. (Fortelius and Kappelman, 1993, 85)
Meanwhile in Spain, about 25 million years ago, another line of giant rhinos was arising which was to culminate in the great elasmotheres of the Pleistocene Ice Age (Kurtén, 1971, 113). It is here that the first evidence for a species of rhinoceros with a single frontal (or forehead) horn, rather than a nasal (or nose) horn appears. As is the case with all Rhinocerotidae, once the genetic basis for a horn exists, the number of horns can vary from one to as many as five (Hillman-Smith & Groves, 1994, 2). This amazing variety is only possible because the “horns” of Rhinoceratidae have no horn cores. In other words, rhino horns are not bony protrusions from the skull, as is the case with bovid horns or cervid antlers, but are composed of bundles of compacted keratin which are attached to a fatty rugose pad of flesh covering a bump on the surface of the skull (fig. 32).

Figure 32. Comparison of deer antler, ox horn and rhino “horn”
In the past, rhino horn was often described as composed of “matted” or “agglutinated” hair. However, according to recent research by Jan Chapman, this is simply not true.
Trying to find a description of the structure of rhinoceros horn which is acceptable to scientists and art historians alike is no easy task, but it is obvious that the term ‘agglutinated hair’ must be laid to rest once and for all. The term ‘agglutinated’ because there is no sticky substance present and the term ‘hair’ because there is no follicle present. So, even though the long hair-like filaments are composed of molecules of the protein keratin as in real hair, the absence of the follicle in rhinoceros horn means that the description ‘hair’ should not be used (Chapman, 1999, 49).
Supplied by a network of blood vessels in the pad, a rhinoceros horn continues to grow, like the hooves of a horse, throughout the lifetime of the animal. Thus if it is wrenched off, wholly or partially, it will eventually regrow (just as human fingernails do).
Fossil “horns” of the Pleistocene woolly rhinoceros Coelodonta Antiquitatis have been recovered from Siberia, where they survive only because they are frozen in the permafrost. They were described for the first time in Western scientific literature in 1769, although it is possible that such “horns” circulated in Europe during mediaeval times (Fortelius, 1983, 125-26). In the West, such fossil rhinoceros horns were mistaken for the huge “claws” (griffes) of the giant mythic raptor known as the gryphon or griffen (Mayor, 2000). In contrast, although its fossilized bones have been recovered, to my knowledge no specimens of the “horn” of the giant unicorn Elasmotherium have ever been published. Because the home territory of the Elasmotherium was not the frozen wastes of Siberia, but the Eurasian steppes, its keratin horn would have been subject to decomposition over time as a natural result of weathering or burial in the earth. In addition, from ancient times the peoples of northeastern Asia used the glue derived from melted rhino horn to increase the elasticity of their composite bows. The other strong possibility is that these great “horns” were believed to be “Dragon’s bones” or “Dragon’s teeth” and ground up for use in Chinese medicine.
In 1934, J. Gunnar Andersson wrote a fascinating account of his discoveries of prehistoric mammal fossils in China, stating:
Most interesting of all the rhinoceros discoveries and perhaps the most significant novelty which China’s Hipparion strata furnished to paleontological science is the curious form which Ringström (1924) described under the name Sinotherium lagrelii. Both the family and the species were new to science. The former name indicates that it was China which contributed this gift to science…. The Sinotherium is a gigantic member of the rhinoceros group, but otherwise shows little relationship to the majority of its forms. The most characteristic feature of the animal is that the enamel of the molars is folded in a manner which reminds us of the equally gigantic Elasmotherium, two species of which are found in soil of the steppes of Southern Russia. There is no doubt that the Sinotherium and Elasmotherium are closely related, even though there is a great difference in age. The Elasmotherium belongs to the latest geological age, the Pleistocene, but the Sinotherium belongs to the transition from the Miocene to the Pliocene. The Sinotherium is the largest of these forms, and there is much to indicate that the whole type had reached its zenith as early as the age of the Hipparion fauna. (Andersson, 1934, 84–85)
Björn Kurtén agrees that the Elasmotherium is “the terminal form of an evolutionary line that has its roots far down in the Tertiary; we have glimpses of it in the Miocene of Spain, the Pliocene of China and the Pleistocene of Russia.…. [It] was a truly gigantic animal, far larger than any living rhinoceros. It had no nose horn, but instead an immense horn on the forehead: it grew to a length of two metres. This animal was thus a veritable unicorn” (Kurtén, 1968, 144-45, emphasis added).
Fossils of elasmotheres have been found in the early and middle Pleistocene layers of northern China, not far from the steppes of southern Russia (Xue and Zhang, 1991, 317). In addition, according to Kurtén, “A few stray finds also indicate that the Elasmotherium made a temporary incursion in central Europe in the Middle Pleistocene” (Kurtén, 1968, 144). However, this was long before the arrival of modern human beings in that part of the world, so there is no possibility that the real unicorn Elasmotherium could have served as a direct source for the mediaeval European myth of the unicorn. The Elasmotherium figures as a giant unicorn with a petrified heart in an Ice Age saga written by the great Danish poet Johannes V. Jensen (Kurtén 1971, 200). But judging from his dates (1873–1950), Jensen must have learned about the existence of the Elasmotherium from the writings of nineteenth century scientists such as Brandt.6
It is true that images of a two-dimensional ox-like male “unicorn” standing beside an altar appear in ancient Indian art on clay seals from the Indus Valley civilization (2600–1900 B.C.E.), and that this impressive beast may have served as the totem of some powerful clan or community (fig. 33).

Figure 22. Modern impression of an Indus valley seal (ROM no number).
But until such time as a three-dimensional image of this animal is found within an archaeological context in India, it is impossible to confirm its status as a true unicorn. In addition, since the Indus Valley script has yet to be deciphered, there is no way to identify it or understand its significance (Edwards, 2000, 126). What is fascinating, however, is how much these images call to mind the Elasmotherium. If, as Kurtén says, “that the Elasmotherium made a temporary incursion in central Europe in the Middle Pleistocene,” then it is also possible that the Elasmotherium could have migrated south-eastward into India, and have survived long enough to provide inspiration for the Indus Valley totem animal. Yet without actual fossil evidence, this must remain sheer speculation. Nevertheless, it is also curious to note that the altars in such images resemble the forms of the ancient Chinese ob pictograph shi 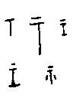 meaning “an object of worship” “an altar” (Hsü, 1996, 882-4). With the collapse of the Indus Valley culture around 1900 B.C.E., these enigmatic images disappeared forever from Indian art.
meaning “an object of worship” “an altar” (Hsü, 1996, 882-4). With the collapse of the Indus Valley culture around 1900 B.C.E., these enigmatic images disappeared forever from Indian art.
Finally, let us return to the real unicorn, for one burning question remains: what did it look like? Unfortunately, Kurtén concludes his discussion of this great beast with the most tantalizing statement of all. “Knowledge of the postcranial skeleton is incomplete” (Kurtén, 1968, 145). Thus although it is possible to reconstruct the appearance of the head of the giant unicorn from fossil remains of its skull, the shape of the rest of its body remains a complete mystery. Nevertheless, several attempts have been made to reconstruct the appearance of the Elasmotherium, including a recent one done at my request by Virginia Mielke (fig. 34).

Figure 34. A reconstruction of the appearance of the Elasmotherium by Virginia Mielke.
On the basis of the evidence presented here, it is no longer possible to deny that the idea of an animal with a single horn projecting from the centre of its forehead, which is the universally accepted definition of a unicorn, is firmly grounded in prehistoric reality. Together with a few other archaic species of giant mammals, the unicorn Elasmotherium survived in the East Asian Refugia until it was probably hunted to extinction in the late Pleistocene period, around 10,000 B.C.E. There is also no doubt that mammalian fossils belonging to the evolutionary line which culminated in the Pleistocene Elasmotherium have been found in Spain and Germany. But in Europe these huge animals died out long before they could be seen by human beings.
In contrast, the survival of the real unicorn in East Asia down into the period of human prehistory, combined with the amazing strength of the Chinese oral tradition, make it perfectly reasonable to suggest that a memory of this fabulous beast survived and found a distant echo in the myth of the Chinese unicorn zhi. As Jordan Paper explains, “Many cultures have strong oral traditions. Where Chinese culture is different is in maintaining the memory capabilities of oral cultures in a highly literate culture due to the logographic nature of written Chinese and the pedagogy involved in learning written Chinese until this century” (Paper, pers. comm. 1997).